Complejo de Golgi: terminología, ocurrencia, morfología, estructura y otros detalles
Complejo de Golgi: terminología, ocurrencia, morfología, estructura, polarización estructural y bioquímica, composición química, funciones y orígenes.
En 1898, mediante un método de tinción con plata, Golgi descubrió una estructura reticular en el citoplasma. El nombre de "aparato de Golgi" generalmente dado a esta estructura es confuso porque sugiere una relación definida con los procesos fisiológicos de la célula.
Hoy parece más apropiado usar el nombre "Sustancia de Golgi" o "Complejo de Golgi" para referirse a este material que tiene propiedades especiales de tinción. Debido a que su índice de refracción es similar al de la matriz, el complejo de Golgi es difícil de observar en las células vivas. El uso del microscopio electrónico ha proporcionado una imagen distinta de este componente, y se ha revelado su estructura sub-microscópica.
Durante años, se pensó que el complejo de Golgi era un artefacto de diversos procedimientos de fijación y tinción. En otras palabras, muchos científicos creían que la estructura observada durante numerosos procedimientos de microscopía y denominada Golgi no existía realmente en la célula viva. Guilliermond (1923), Parat (1927), Walker y A lien (1921), surgieron dudas con respecto a la existencia del complejo de Golgi.
Terminología del complejo de Golgi:
Holmgren se refirió al complejo de Golgi como trofospongio (Cajal lo llamó canales de Golgi-Holmgren). Baker utilizó el término lipocondria debido al presunto contenido de lípidos. El término Complejo de Dalton se dio después del nombre de su observador Dalton en 1952. Sjostrand propuso el término de cito-membranas para el sistema de Golgi.
Sosa ha sugerido la siguiente nomenclatura para el complejo de Golgi:
1. Golgiokinesis :
División del aparato de Golgi durante la división nuclear.
2. Golgiosomas :
Los corpúsculos producidos por la Golgiogénesis se denominan Golgiosomas y se describen como material de Golgi en invertebrados.
3. Golgiolisis :
Proceso de disolución del aparato de Golgi.
4. Gogiorrhexis :
Fragmentación sobre el aparato de Golgi.
5. Golegiogénesis :
Formación y diferenciación del cuerpo de Golgi durante el desarrollo embrionario.
6. Golgio-citoarquitectura :
Estudio de la estructura celular en relación con el aparato de Golgi.
Ocurrencia:
El complejo de Golgi se produce en todas las células, excepto en las células procarióticas (es decir, micoplasma, bacterias y algas verdes azules) y en células eucariotas de ciertos hongos, células espermáticas de briófitos y pteridofitas, células de tubos de tamiz maduro de plantas y esperma maduro y sangre roja. Células de los animales.
Morfología del complejo de Golgi:
La morfología del complejo de Golgi varía de una célula a otra en función del tipo de célula en que se encuentran. Se han observado dos formas de complejo de Golgi.

1. Forma localizada :
En las células polarizadas de vertebrados (que tienen base y ápice), el complejo de Golgi se produce individualmente y ocupa una posición fija. Se encuentra entre el núcleo y el polo secretor. Esto se puede ver mejor en las células tiroideas, en las células exocrinas del páncreas y en las células mucosas del intestino.
2. Forma difusa :
En algunas células especializadas de vertebrados (células nerviosas y células del hígado), en la mayoría de las células vegetales y en las células de los invertebrados, varias unidades del complejo de Golgi se encuentran dispersas junto con los elementos del retículo endoplásmico. Cada unidad se llama un dictyosome. En las células hepáticas se producen hasta 50 dictyosomas por célula y en ciertas células vegetales su número puede llegar hasta cientos.
Forma :
La forma del complejo de Golgi es bastante variable en diferentes tipos de células somáticas de animales. Incluso en la misma célula hay variaciones en diferentes etapas funcionales. La forma es, sin embargo, constante con cada tipo de célula. Su forma varía de una masa compacta a una red filamentosa dispersa.
Número:
El número de acumulaciones de Golgi por celda varía enormemente, dependiendo del tipo de celda, desde solo una hasta cientos. Hay una sola grande en algunas celdas, mientras que en el caso de Paramoeba hay dos. En Stereomyxa (una especie de Amoeba) hay muchos complejos de Golgi. Las células nerviosas, las células hepáticas y la mayoría de las células vegetales también tienen complejos de Golgi múltiples, con aproximadamente 50 células hepáticas.
En los rizoides algales se ven más de 25, 000 complejos de Golgi (Sievers 1965). El complejo de Golgi puede incluso dar cuenta de una gran fracción del volumen celular en algunas células especializadas. Un ejemplo es la célula caliciforme del epitelio intestinal, que secreta moco en el intestino; Las glicoproteínas en el moco están glicosiladas principalmente en el complejo de Golgi.
Tamaño:
El tamaño es igualmente variable. Es grande en las células nerviosas y glandulares y pequeño en las células musculares. En general, el complejo de Golgi está bien desarrollado mientras que la célula está en estado activo. Cuando la célula envejece, el complejo disminuye progresivamente en tamaño y desaparece.
Posición:
La posición del complejo de Golgi es relativamente fija para cada tipo de célula. En las células de origen ectodérmico, el complejo de Golgi está polarizado desde el momento del estado embrionario entre el núcleo y la periferia (Cajal, 1914). En las células exocrinas secretoras que tienen en general una polarización típica, el complejo de Golgi se encuentra entre el núcleo y el polo secretor.
En las glándulas endocrinas, la polaridad de este órganoide es variable, excepto en la tiroides, donde está orientada hacia el centro del folículo. En las células más jóvenes y, a menudo, en las más antiguas, se encuentra con mayor frecuencia en un lado del núcleo, pero en ciertos casos puede rodearlo completamente. En las células ganglionares del ratón la posición es perinuclear.
Estructura detallada del complejo de Golgi:
Dalton y Félix (1954) describieron el complejo de Golgi en el epidídimo de la rata después de tomar las primeras micrografías electrónicas.
La siguiente descripción del complejo de Golgi es un compuesto basado en el trabajo de varios autores:
1. Cisternas :
Las cisternas o sacículos son similares a la superficie lisa ER, y aparecen en sección como pilas de sacos delimitados por membranas muy espaciados. El número de sacículos varía de 4 a 8 en la mayoría de los tipos de células animales y vegetales. En Euglena, el número puede llegar hasta 20.
La membrana de las cápsulas tiene un grosor aproximado de 60 a 70 A °, que encierra una cavidad de alrededor de 150 A ° de ancho, cuyos bordes a menudo están dilatados. Según la mayoría de los autores, hay dos caras bien definidas de las cisternas, es decir, convexas y cóncavas; esta última generalmente se conoce como cara madura o en formación o distal y se supone que el lado convexo es la cara inmadura o excitada o proximal, los cistéranos que se encuentran en una matriz paralela están separados entre sí por un espacio de aproximadamente 200 a 300 A 0 .
Aún no se sabe qué es lo que los mantiene unidos, pero en pocas células se ve una capa delgada de electrones opacos, a veces densos, entre las cápsulas, que en ciertas regiones son más prominentes a las que Amos y Grimstone (1968) aplicaron el término nodos. Mollenhauer et al., (1973) exploraron con cierto detalle elementos y placas intercisternales en ciertas plantas del complejo de Golgi.

2. Los túbulos :
Del área periférica de las cisternas surge una red plana y anastomosada de túbulos de 300 a 500 A de diámetro. Clowes y Juniper (1969) han comparado esta red tubular con un disco de encaje.
3. Vesículas :
Las vesículas son pequeños sacos en forma de gotitas que permanecen unidos a los túbulos en la periferia de las cisternas.
Son de los siguientes dos tipos:
(a) Vesículas lisas :
Las vesículas lisas son de 20 a 80 µ de diámetro. Contienen material secretor (que a menudo se llaman vesículas secretoras) y se desprenden de los extremos de los túbulos cisternales dentro de la red. A menudo, más de una conexión de túbulos, y presumiblemente se llena, una vesícula de formación única.
(b) Vesículas recubiertas :
Las vesículas recubiertas son protuberancias esféricas, de aproximadamente 50 µm de diámetro y con una superficie rugosa. Se encuentran en la periferia del orgánulo, generalmente en los extremos de los túbulos individuales y son morfológicamente muy distintos de las vesículas secretoras. Su función es desconocida.
4. Vacuolas de Golgian :
Estos son grandes sacos redondos presentes en la cara de maduración de Golgi. Estos se forman ya sea por las cisternas expandidas o por la fusión de vesículas secretoras. Las vacuolas están llenas de alguna sustancia amorfa o granular.
El complejo de Golgi está polarizado estructural y bioquímicamente:
El complejo de Golgi tiene dos caras distintas: una cara cis o en formación y una cara trans o en proceso de maduración. La cara cis está estrechamente asociada con una parte de transición suave de la ER en bruto. En las células secretoras, la cara trans es la cara más cercana a la membrana plasmática: aquí, las vesículas secretoras grandes se encuentran exclusivamente en asociación con la cara trans de una pila de Golgi, y la membrana de una vesícula secretora en formación a menudo es continua con la de La cara trans de la última cisterna ("trans-most").
En contraste, las pequeñas vesículas de Golgi están localizadas de manera más uniforme a lo largo de la pila, se piensa que las proteínas ingresan a una pila de Golgi desde la sala de emergencia en el lado cis y salen para múltiples destinos en la parte trans; sin embargo, no se conoce ni su ruta exacta a través del complejo de Golgi ni cómo viajan de cisterna a cisterna a lo largo de cada pila.
Las dos caras del complejo de Golgi son bioquímicamente distintas. Por ejemplo, una variación en el grosor de las membranas de Golgi puede detectarse a través de la pila en ciertos casos, siendo las del lado cis más delgadas (tipo ER) y las del lado trans más gruesas (como la membrana plasmática).
Más notables son los resultados obtenidos cuando se usan ciertas pruebas histoquímicas junto con microscopía electrónica para localizar proteínas particulares dentro del complejo de Golgi. Algunas de estas pruebas revelan enzimas unidas a la membrana, actividades que muestran una polaridad distinta en su localización dentro de la pila de Golgi.
Un hallazgo bioquímico particularmente intrigante fue el descubrimiento de que las enzimas lisosomales, como la fosfatasa ácida, se concentran con la cisterna más trans de la pila de Golgi y dentro de algunas de las vesículas recubiertas cercanas. Esto sugiere que las vesículas específicas que salen para los lisosomas se ensamblan en esta región.
Las proteínas secretoras se encuentran por métodos histoquímicos en todas las cisternas apiladas, a pesar de que las grandes vesículas secretoras en las que estos productos se concentran y se asocian solo con la cisterna de Golgi más trans.
Composición química:
Con respecto a la composición química del complejo de Golgi, se ha demostrado que las siguientes sustancias están presentes:
1. Fosfolípidos :
La composición de fosfolípidos de las membranas de Golgi es intermedia entre las membranas endoplásmicas y las membranas plasmáticas.
2. Proteínas y enzimas :
El complejo de Golgi de diferentes células vegetales y animales muestra grandes variaciones en los contenidos de proteínas y enzimas. Algunas de las enzimas son ADPasa, ATPasa, NADPH citocromo-C-reductasa, glicosy1 transferasa, galactosy1 transferasa, pirofosfato de tiamina, etc.
3. Hidratos de carbono :
Tanto las células vegetales como las animales tienen algunos componentes de carbohidratos comunes, como glucosarina, galactosa, glucosa, manosa y fructosa. La planta de Golgi carece de ácido siálico, pero se produce en alta concentración en el hígado de rata. Algunos carbohidratos como la xilulosa y la arabinosa están presentes solo en las células de las plantas.
4. Vitamina С :
Tomitte ha demostrado la fracción de vitamina С almacenada en el complejo de Golgi. Según él, el complejo de Golgi almacena vitamina С y la libera lentamente en el citoplasma en cantidad suficiente para evitar la oxidación de los productos celulares.

Funciones del complejo de Golgi:
1. Formación de acrosoma durante la espermiogénesis :
Durante la maduración de los espermatozoides, el complejo de Golgi desempeña un papel en la formación del acrosoma (Burgos y Fawcett, 1955).

En las primeras etapas, el Golgi aparece como un cuerpo esférico, que comprende cisternas dispuestas en pilas paralelas y numerosas vesículas pequeñas. Lo último siempre arrancado de las cisternas. A medida que avanza el desarrollo, el complejo de Golgi se vuelve de forma irregular y los vaculoles grandes se forman por dilataciones de los sacos cisternales.
En el centro de estas vacuolas grandes o vacuolas está presente un gránulo denso, el gránulo proacrosómico. Este gránulo que se deriva del complejo de Golgi continúa creciendo dentro de la vacuola por un proceso conocido como acreción. Esta vacuola y gránulo se aproxima al polo anterior de la membrana nuclear, constituyendo gránulo acrosomal.
Con el alargamiento de la espermátida, la vesícula acrosomal se extiende sobre la superficie nuclear y finalmente se colapsa con la membrana nuclear, formando el material de la tapa. El gránulo acrosomal se convierte en el acrosoma que se encuentra en el vértice del núcleo y aparentemente comprende ciertas enzimas involucradas en el proceso de fertilización.
2. Síntesis y secreción de polisacáridos:
Los estudios en autómatas mediante autorradiografía y microscopía electrónica han establecido la interrelación entre la síntesis de proteínas, la adición de carbohidratos y la sulfatación. Las células caliciformes del colon producen mucígenos. Este material secretor contiene una gran proporción de carbohidratos.
El complejo de Golgi se encuentra justo encima del núcleo. Hacia la superficie libre de la célula se están agrandando gradualmente los gránulos mucigénicos. Las cisternas proximales del complejo de Golgi no muestran hinchazón, pero a cierta distancia a través de la pila, las cisternas distales se convierten repentinamente en gránulos mucigénicos. Las cisternas distales se convierten continuamente en gránulos de mucógeno cada 2-4 minutos. Las nuevas cisternas proximales se forman en compensación.
3. Papel en la secreción:
Se considera que el complejo de Golgi desempeña algún papel en la función secretora de una célula. Pero la cuestión es que están secretando o sintetizando algunas sustancias o son simplemente un almacén en el que los productos secretores que se secretan en otro lugar de la célula, simplemente se almacenan y se concentran.


A partir de los estudios de Palade et al. 1962 este ciclo secretor ahora está bien definido e incluye cuatro pasos en el caso de células acinares pancreáticas y son:
(i) La incorporación de aminoácidos a la proteína en la superficie del retículo endoplásmico rugoso.
(ii) Transferencia de estas proteínas secretoras nacientes a las cisternas del retículo endoplásmico rugoso.
(iii) El transporte intracelular de estas proteínas al complejo de Golgi.
(iv) La migración de gránulos de zimógeno hacia el vértice de la célula donde se descargan en lúmenes.
4. Papel del cuerpo de Golgi en la ovogénesis :
Srivastava (1965) ha hecho una breve reseña del complejo de Golgi durante la ovogénesis. Según Afzelius (1956), el complejo de Golgi de un huevo de erizo de mar, como se ve bajo el microscopio electrónico, consiste en pilas de láminas que forman paredes de bolsas planas, que ocasionalmente pueden estar hinchadas.
Hay algunas indicaciones de divisiones transversales de estos cuerpos. Sotelo (1959) y Sotelo y Porter (1959) describieron el complejo de Golgi en el óvulo de rata como se observa en el microscopio electrónico y encontraron una localización nuclear de este orgánulo en los primeros ovocitos.
En la siguiente etapa, estos se resuelven en fragmentos y en la tercera etapa, estos se mueven hacia la corteza. En todos estos casos, su estructura sigue siendo de conjuntos muy compactos de esbeltos, perfiles dobles (sacos aplanados) y vesículas esféricas.
En los primeros ovocitos el complejo está organizado de forma compacta. En etapas posteriores, se encuentran dispersos haces de perfiles, rodeados por pequeñas vesículas, dispersos en la zona cortical. En los primeros ovocitos, el complejo de Golgi y el centrosoma están estrechamente asociados.
5. Absorción de compuestos :
Hirsch et al., Han descubierto que cuando el azúcar de hierro se alimenta a un animal, el hierro se absorbe en el complejo de Golgi (Kedrowsky). Van Teel ha demostrado que los sistemas de Golgi también absorben compuestos de cobre y oro. Kedrowsky ha demostrado que el complejo de Golgi de Opalina puede absorber bismutosa (compuesto de albúmina y bismuto) y protargol (compuesto de albúmina y plata). Por lo tanto, Kirkman y Severinghaus afirman que el complejo de Golgi actúa como una membrana de condensación para la concentración de productos en gotitas o gránulos.
6. Formación de la pared celular de la planta :
La pared celular de las plantas está formada por fibrillas que contienen predominantemente polisacáridos, junto con algunos lípidos y proteínas. Durante la citocinesis, se forma una placa celular entre los dos núcleos hijas, y tiene alrededor una membrana que luego se convierte en la membrana plasmática de las células hijas. Existe una clara evidencia de que los polisacáridos se forman en el complejo de Golgi y se transfieren a la nueva pared celular que se establece mientras las células aún están creciendo.

El complejo de Golgi también contribuye con sustancias como pectinas y hemicelulosas, que forman la matriz de la placa celular que separa las membranas plasmáticas.
7. Formación de cristales intracelulares :
En el isópodo marino, Limnoria lingmorum, que es una forma de excavación, están presentes las glándulas medias, cuyas células están formadas por cristales. Estos varían hasta 30 A ° de longitud y 15 A ° de espesor. Se ha demostrado que estos cristales están formados por el complejo de Golgi y se sabe que contienen proteínas y hierro. No tienen membrana envolvente y suelen ser de forma esferoidal. Están preocupados por la actividad secretora.
8. Formación de gotitas de proteína de leche :
En la glándula mamaria lactante de ratones se producen gotitas de proteínas que están relacionadas con el complejo de Golgi. Estas gotitas generalmente se abren a la superficie celular mediante la fusión de su membrana envolvente con la membrana plasmática.
9. Formación de lisosomas y vacuolas :
Los lisosomas primarios se forman a partir de las membranas de Golgi de la misma manera que las vesículas secretoras. Existe buena evidencia de que los dictyosomas acumulan enzimas hidrolíticas en sus regiones más maduras. Se ha encontrado que algunas vacuolas en células vegetales contienen pequeñas cantidades de enzimas hidrolíticas y se presume que éstas se derivan del complejo de Golgi.
10. Formación de pigmentos:
En muchos tumores de mamíferos y células cancerosas, el complejo de Golgi se ha descrito como el lugar de origen de los gránulos de pigmento (melanina).
11. Regulación del balance de fluidos :
Se ha sugerido una homología entre el complejo de Golgi y la vacuola contráctil del Metazoa inferior y los Protozoos. La vacuola contráctil expulsa el agua sobrante de la célula. En ciertos Protozoos, el complejo de Golgi también se ocupa de la regulación del balance de líquidos.
Origen del Complejo de Golgi:
Se han propuesto tres fuentes diferentes de las que puede surgir el nuevo complejo de Golgi:
1. Desde el retículo endoplásmico :
Essner y Novikоff (1962) y Beams y Kessel (1968) han propuesto que las cisternas de Golgi surjan de la sala de emergencias. El retículo endoplásmico rugoso después de sintetizar proteínas específicas pierde ribosomas y se convierte en ER suave. Pequeñas vesículas transitorias se desprenden de la ER suave. Estos migran a dictyosome.
Al llegar a la cara de formación de dictyosome estos se fusionan para formar nuevas cisternas y así contribuir a su crecimiento. Mediante la fusión de estas vesículas, se forman nuevas cisternas continuamente en la cara que se forma y en la cara de maduración las cisternas antiguas se descomponen en vesículas secretoras. Así, Golgi exhibe un fenómeno de flujo membranoso '.
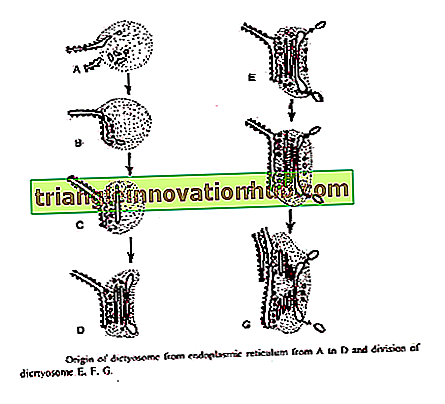

Bouch (1965) describió el origen de Golgi a partir de la membrana externa de la envoltura nuclear en algas pardas. Las vesículas se separan de la membrana nuclear externa que se fusiona para formar cisternas en la cara de formación de dictyosome.
La presencia de zonas de exclusión en relación con la ER suave o la membrana nuclear, la aparición de zonas de exclusión en semillas latentes de plantas superiores y la formación de diciosoma de estas zonas en germinación de semillas proporcionan evidencia en apoyo de las dos teorías anteriores sobre el origen de dictyosome.
3. Por la división de dictyosome preexistente :
Se ha observado que durante la división celular tanto en plantas como en animales, el número de dictyosomes aumenta y el número de dictyosomes en cada célula hija justo después de la división es casi igual al número en la célula madre antes de la división, de latas y otras. En las observaciones sobre las células en división, se ha supuesto que los diciosomas también se dividen durante la división celular.